
The great Dutch biologist Niko Tinbergen, first described the highly stereotypic courtship behaviour of the Grayling, Hipparchia semele (Linnaeus 1758), a butterfly which was common on a dune area in the center of the Netherlands, Hulshorst sands – on the shores of the Zuider Zee – where he was carrying out research on digger wasps. The complex courtship sequence carried out by the male Grayling involves a series of moves and maneuvers, which if successful, end in copulation (See description in Link 1). The key element of the sequence he called ‘bowing‘ and this is also exhibited by the Rock Grayling, as described below. Tinbergen describes the behaviour of the male as follows:
After alighting near the female, he walked round until he faced her. The with curiously jerky movement, he raised his forewings step by step, quivering them, and with one final forward jolt hit the female with them. All the time the wings were kept almost or completely folded.
Tinbergen called Grayling butterflies ‘bark with wings‘ in a chapter in his little book entitled Curious Naturalists (Tinbergen, 1984) and their tree bark-like patterning gives them remarkably good camouflage when resting on tree trunks (below). They are however, still apparent in profile, especially if lit by the sun as here.

Rock Graylings (Hipparchia hermione) have a patchy distribution in Europe, occurring in parts of Spain, southern France, Italy, southern Finland as well as in central Europe. N.B. The species was formerly called H. alcyone, but that name was reportedly found to be preceded by H. hermione.(2, 3) The Rock Grayling is difficult to distinguish from the closely related Woodland Grayling (H. fagi), but as far as I am aware – and this confirmed by maps in at least three guides – the Rock Grayling is the only one of these two species to occur in NW Spain.

Working in the Central Appennines (Rieti, Italy), Manuela Pinzari of the Department of Biology, University of Rome “Tor Vergata”, showed that the general pattern of sexual behaviour in the Tree Grayling (H. statilinus) was almost identical to that of the Grayling (H. semele) (Pinzari, 2009). The only differences appeared to be in the ‘presence/absence, order and performance of steps’ in the courtship sequences.

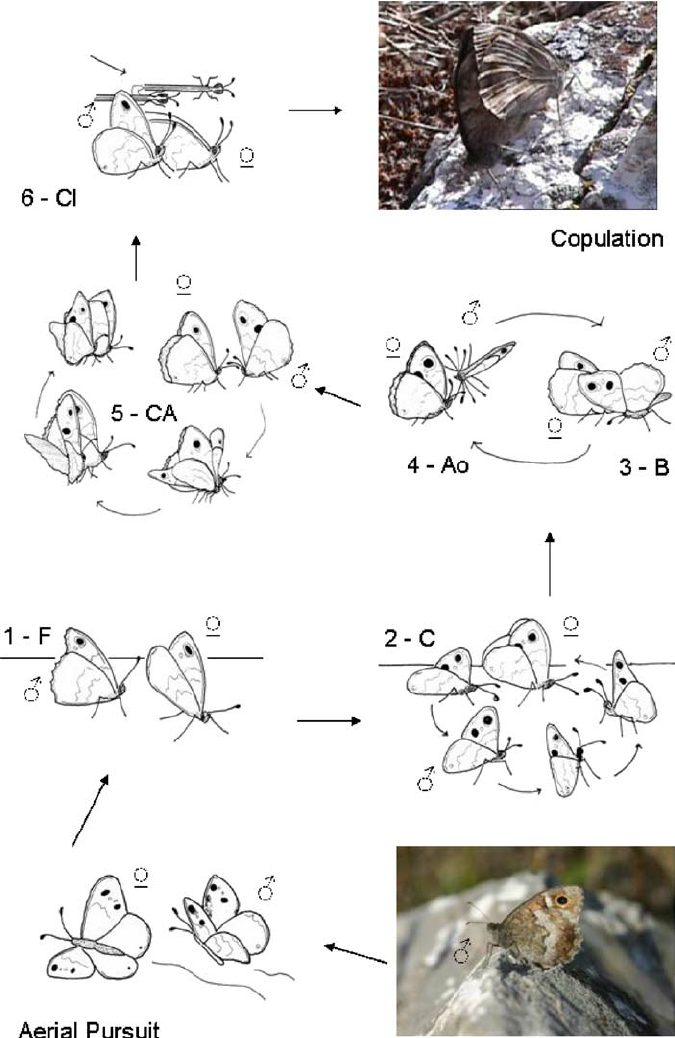
Building on the pioneering studies by Tinbergen (1941, 1972), Pinzari (2009) describes six patterns of behaviour in the courtship sequences of both the Grayling (H. semele) and the Tree Grayling (H. statilinus). I have taken the liberty of summarising these as follows:
- – Fanning. The male lands on the ground behind the female with his wings closed, but revealing the large post-discal ocellus. The fore-wings are rapidly opened and closed in a rhythmical, vibration-like motion.
- Circling. The male walks around the female – in a semicircular track – so he ends up facing her head on, whilst fluttering his wings.
- Bowing. The male starts bowing even before he is in front of the female. Initially, his wings are closed; he then rotates his body forward on the sagittal axis, levering upwards on the posterior legs; whilst his head is brought close to the ground. Simultaneously, both wings open and close with great speed. Bowing ends when the male closes his fore-wings and moves them back to the initial position, along with the body. During very deep bowings, the male kneels on the ground. While bowing, the male can take hold of the female fore-wings and antennae, letting them slide between his fore-wings.
- Antenna orientation. Immediately after bowing, the male antennae are waved back and forth very rapidly (this can only be seen clearly in slow motion it seems).
- Copulation attempt. This resembles an unfinished bowing movement with an “embrace”. The male remains bowed forward with the wings open and in this bowed position, walks towards the side of the female, then behind her, until they both face the same direction.
- Clasping. The male attempts copulation by flexing his abdomen sideways and making contact with the female genitalia.
These behaviours are beautifully illustrated in a series of sketches in Fig. 2 in Pinzari (2009) (below).
One interesting finding of the study by Pinzari (2009) was that it revealed that, whether or not the courtship progressed as far as copulation, largely depended on the female. In short, it appears that the whole sequence was often interrupted and restarted again at an earlier point in the cycle. The female communicates her availability – or otherwise – to the male, by body language. For example, she may just become inactive and fail to respond, and she has other ways – flapping and opening her wings – of rejecting or discouraging persistent males (see (Pinzari & Sbordoni, 2013). Pheromones (scents) produced by the male also play a major role in the courtship and the bowing behaviour is thought to be a way of exposing the female to these scents and thereby stimulating her into a state of receptivity. This process may also allow the female to evaluate the quality of the male and if he is not to her liking, she can break off the affair!
During Bowing, ‘the male bashes and strokes the female antennae between his forewings on the androconial scales revealing the importance of scent stimuli in mate communication’ (Pinzari & Sbordoni, 2013).

Manuela Pinzari, together with co-worker, V. Sbordoni, then went on to investigate the courtship behaviour of a subspecies of the Rock Grayling (H. hermione genava), with the aim of investigating whether there were any differences in courtship behaviour between this species and the Woodland Grayling, which might lead to reproductive isolation between the two species (Pinzari & Sbordoni, 2013). It turned out that although the overall pattern of sexual behaviour was similar in the two species, there were some significant differences with regard to the bowing behaviour. In summary, these detailed and I think, equally pioneering studies showed that although the courtship sequence in the three Grayling species (H. fagi, H. hermione and H. semele) is similar, there are differences in terms of when the steps are carried out and in what order (Pinzari & Sbordoni, 2013).
A key finding of these studies is that when interrupted, a male can re-start the courtship from any step of the sequence. On only about 10% of the time was a single courtship sequence followed by copulation. In other words, the males had to court the females repeatedly and this means that one is very unlikely to see the whole courtship sequence, from start to copulation, in the field. The Bowing and Antenna orientation behaviours could be repeated many times in a row (up to 25 times in the Rock Grayling, H. hermione).

So the take home message, if there is one, is that certain relatively short sequences of behaviour – of the sort shown below – could be repeated numerous times, perhaps with interruptions by other males, before reaching a final conclusion.
Although I had heard about the bowing behaviour of Grayling butterflies, I was unaware of the detailed studies by Pinzari and co-workers when I took these photographs. I came across many Rock Grayling butterflies defending territories in Galicia, Spain, in August 2017, and I was struck by the behaviour I witnessed on a couple of occasions. As Tinbergen described (quotation above), the bowing behaviour is quite a jolt. It looked to me at first, before I had worked out what was going on, like a butterfly head-butt! Anyway, I think I managed to capture this behaviour in a photograph, a couple of times in the field. See Photographs # 5 and 10 (below).
The sequence of photographs below involved the same pair, although they were interrupted at one stage by another individual (probably another male) (See #9 below). After they all fly up into the gorse bush it is difficult to determine whether the original pair continued, or whether the first male was usurped by the interloper! Assuming that it was the original pair, they then flew back down to the ground and continued their courtship. Given, based on the work of Pinzari & Sbordoni (2013), that the Bowing and Antenna orientation behaviours could be repeated up to 25 times in a row, such interactions could probably go on for hours! Unfortunately, I did not witness any coupling in this species. It could be that they fly up into the trees whilst in copula.
Courtship Sequence captured in photos













Populations of Rock Graylings appear very healthy in Galicia, and assuredly in other regions of Spain; there were large numbers sun basking on unpaved tracks in the hilly farmlands, but their status in northern Europe is less rosy. “During the last decades, Hipparchia alcyone has experienced such a strong decline in the Federal Republic of Germany that it is presently threatened by extinction” (Kwast & Sobczyk, 2000).
Links
- http://www.learnaboutbutterflies.com/Britain%20-%20Hipparchia%20semele.htm
- http://www.eurobutterflies.com/sp/hermione.php
- http://www.butterfliesoffrance.com/html/Hipparchia%20alcyone.htm
- https://www.gbif.org/species/1918851
- http://www.inaturalist.org/taxa/362064-Hipparchia-alcyone
- http://www.guypadfield.com/rockgrayling.html
- http://www.luontoportti.com/suomi/en/perhoset/rock-grayling
References
Kwast, E., & Sobczyk, T. (2000). Ecological requirements and distribution of Hipparchia alcyone (Denis & Schiffermüller, 1775) in the Federal Republic of Germany (Lep., Satyridae). Entomologische Nachrichten und Berichte, 44(2), 89-99.
Pinzari, M. (2009). A comparative analysis of mating recognition signals in graylings: Hipparchia statilinus vs. H. semele (Lepidoptera: Nymphalidae, Satyrinae). Journal of insect behavior, 22(3), 227-244.
Pinzari, M., & Sbordoni, V. (2013). Species and mate recognition in two sympatric Grayling butterflies: Hipparchia fagi and H. hermione genava (Lepidoptera). Ethology Ecology & Evolution, 25(1), 28-51.
Tinbergen N (1941) Ethologische Beobachtungen am Samtfalter, Satyrus semele L. Journal of Fuer Ornithologie 89(3):133–144.
Tinbergen N (1972) The courtship of the Grayling Hipparchia (=Satyrus) semele (L.). In: London. Allen & Unwin (ed) The animal in its world, field studies. vol. I, pp 147–249.
Tinbergen, N. (1984). Curious naturalists. Univ of Massachusetts Press.

[…] Courtship behaviour of the Grayling butterfly via Ray Cannon […]
[…] is elaborate and consists of a series of complex courtship sequences (See previous blog on the courtship behaviour of the Rock Grayling (Hipparchia hermione). Indeed, as one author – Matthew Oates – eloquently put it in a […]
[…] is evaluating his quality as a potential mate during this process. Graylings have a particularly elaborate courtship, with a repertoire of moves and motions, including the famous ‘bowing’ behaviour […]
[…] I do not have photographs of this behaviour in Graylings (yet!), but I have photographed it in the Rock Grayling. Tinbergen described it as follows: the male ‘captures the female’s antennae between his […]
[…] semele) and rock graylings (Hipparchia hermione) engage in courtship bowing, as described in another blog of mine. The bowing behaviour is so distinctive (see below), that once you have seen it, you never forget […]
[…] have been lucky enough to capture, is courtship bowing in Rock Graylings (below), as I described in this blog, called Rock (and roll!) Graylings! Although the courtship behaviour in these different Grayling […]