
The Common Blue is a gorgeous little butterfly. Although the bright blue male is flashy and showy, it’s the females I like best, with their subtle variations of blue and brown. Why these variations – blue to brown – persist and vary has puzzled scientists for many years; here I summarise some recent research on pigments and structural colours, and speculate on how it might work!
The colour of the upperwings of females varies from almost completely brown in southern England to predominantly blue in western Ireland and Scotland. (Butterfly Conserrvation)
The Common Blue (Polyommatus icarus) is one of the commonest and widespread lycaenid butterflies in Europe, although declines in distribution and populations have been reported in recent years from a number of countries, including Spain – where some of these photos were taken – and the United Kingdom (below).

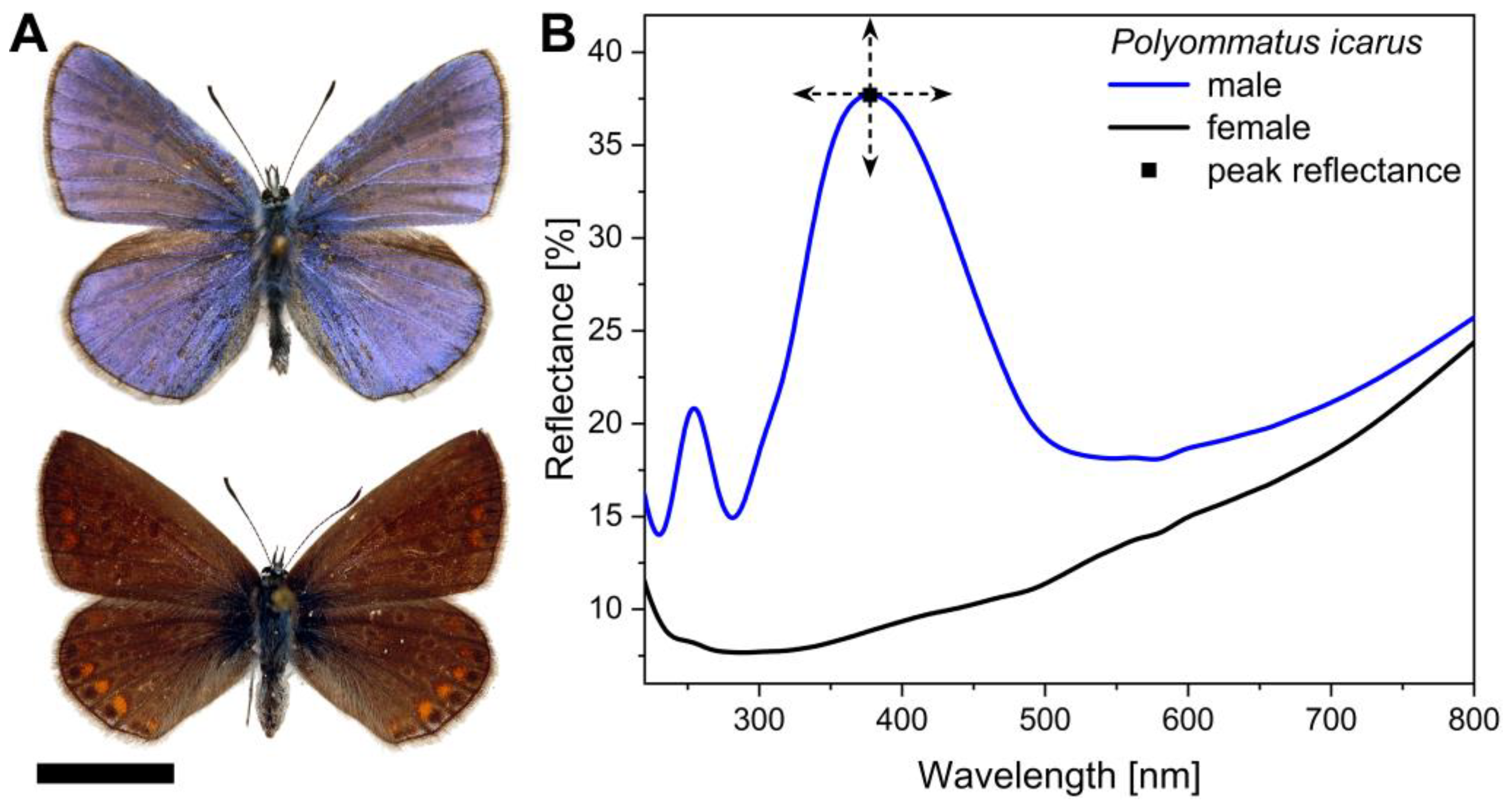
Like many lycaenids, Polyommatus icarus exhibits sexual dichromatism; a fancy name for where the males and females are differently coloured. In this case, the dorsal (upper) wing surfaces of the males are a bright, UV-reflecting blue colour as a result of nano-structures: photonic crystals which reflect light. In P. icarus males cover scales on the wing surface mainly reflect short wavelengths (UV/blue colours). The brown colours of the females wings on the other hand, are a result of pigments, light absorbing molecules such as melanin, which is present in the underlying basal wing scales (De Keyser et al., 2015).

Female Common Blue’s are – like the closely related Adonis Blue – are highly variable in terms of coloration. They show a continuous variation from blue to brown as a result of the density of the blue-reflective wing scales, although not necessarily at one site. For example, female Common Blues in Western Ireland and Scotland are said to be almost completely blue, according to Steve Cheshire on his informative site, as are females in more southerly, hotter sites in Europe such as Turkey and Morocco. There is some evidence for an environmental influence on the so-called blue suffusion, with both hot and cold temperature stress generating the bluer females (Kertész et al., 2017). In experiments, prolonged cooling induced the appearance of the blue scales on the dorsal wing surface of the females. Interestingly, the blue scales on the females that appeared as a result of cooling stress in the experiments, were not as structurally sound as those in the males: they ‘had fewer layers and a more disordered nanoarchitecture’ than the blue scales of the males (Kertész et al., 2017). So it is tempting to conclude that they may not function as well as the UV-reflecting blue scales of the male.

The conspicuous blue coloration of all male of lycaenid butterflies is generated by photonic nanoarchitecture – i.e. nanoscale architecture – on their dorsal wing surfaces (below). The structural blue colour of the male has a characteristic reflectance peak at a wavelength of around 400 nm, while the brown females lack this.
The structural colours of the polyommatine lycaenid butterflies are generated by a particular type of “pepper-pot” photonic nanoarchitectures (see Piszter et al., 2023).

Common Blues are called dorsal baskers! Not a rude word, but a term for sitting with their wings held open, warming up in the sun (above). You might think that the darker brown females, with their dark melanin pigments, would warm up faster than the shiny males? But it turns out that variation in wing melanisation, both within and between sexes, had little or no effect on how quickly the butterflies were able to warm up (De Keyser et al., 2015). This would seem to explain why you don’t just get more browner females as you go north. So what’s the advantage in being brown we might ask? Predominantly brown females appear to be commonest in Central European countries at the centre of range. One theory is that the fact that the bluest females are to be found at the range margins, is a relic property from when the species survived in refuges during the ice ages. The blue colour may be associated with some other survival feature which gets switched on by cold (Kertész et al., 2017). All down to an ancient adaptation to survive the ice-ages?!

One very interesting recent piece of research has revealed that Polyommatus icarus male butterflies see colours in the green part of the light spectrum (extending up to 560nm), but remarkably they cannot distinguish colors in the red range (570–640nm) of the spectrum, for example they cannot not distinguish yellow from red (Sison-Mangus, 2008). They are colour blind! Or at least partially colour blind, as they can distinguish between yellow and blue! So the lovely colourful patterns on the dorsal wings of the females – the submarginal orange or red crescents (called ‘lunulation’) – are apparently invisible to the male. Instead, his eyes are said to be ‘perfectly tuned to the coloration displayed by male dorsal wing surfaces’ (below), in other words to blue (Kertész et al., 2017).

So the males are usually the most conspicuous sex in terms of appearance and generate a stronger optical signal than the female. The male vision is well-tuned to see these blue colours, which is useful for interactions with other males, but what attracts them to the female? It may in part, be the bright blue scales, but it has been discovered by researchers in Germany that it is also the presence of flavenoids in their wings that attracts males (Burghardt et al., 2000; 2001). Flavonoids are plant pigments which these butterflies obtain when eating their host plants (Medicago sativa, Trifolium repens, T. pratense) as larvae. Females tend to have much more of these UV-absorbing pigments than males and it greatly increases their attractiveness to mate-searching males. Exactly what effect they have on the ‘colouration’ of the females wings is difficult to imagine, because unlike us, butterflies can see ultraviolet light. So whilst the flavenoid-rich females may look the same to us, their UV-absorbing properties make them particularly conspicuous to the males. So the females have both UV-reflecting (blue scales) and UV-absorbing (flavenoid pigments) on the wings, and perhaps it is the balance between these two extraordinary features which determines their attractiveness to males?

It has been pointed out before that the bluest of the female phenotypes (or forms) are more male-like in appearance, and therefore might be favoured by sexual selection (Brakefield & Shreeve, 1992). The bluer females may get noticed first, and courted and mated before their darker sisters. On the other hand, they might also get eaten first, because they look so bright and are fairly sedentary! The more browner female forms might have better survival rates because of their cryptic colouration, but they might never get noticed and fail to mate. In other words, as Brakefield and Shreeve pointed out in their 1992 chapter, there could be a balance of selective forces – sexual (male choice) and natural selection (visual predation) – involved, which would explain why there is an ‘underlying genetic variation’. But we now know that this is not the whole story because there are also variations in UV-absorbing pigments which similarly influence female attractiveness.

It is hard to know exactly what is going on; more research is always needed! But it is tempting to speculate that both the blue scales and the flavenoid pigments – so diligently accumulated by the larval stages – increase the conspicuousness and attractiveness of the female butterflies. The more attractive individuals, will – like Cinderella at the ball! – get chosen first, but they run the risk of being gobbled up by a hungry bird before that can happen! The shyer, less gaudy females, with their dark brown wings and perhaps only a fine dusting of blue scales (below), lurk in the background and avoid being eaten, but may also get missed by all but the most ‘eagle-eyed’ males!

The balance of costs and benefits, from the blue end to the brown end of the spectrum, may be constantly shifting. It all goes to show that you don’t need to go to a tropical rain forest to find butterflies with fascinating biology and many unanswered questions! There is so much to learn about our own native species.

All images taken by Raymond JC Cannon. N.B Some of these were taken back in 2017, at a site in NW Spain (Galicia) where they appear to be doing quite well: Morouzos Beach (or Praia de Morouzous-Cabalar), a protected dune land ecosystem on the Ria Ortigueira, Galicia, Spain. Other images were taken this year (2023), closer to home, in Bedfordshire.
References
Artemyeva, E. A. Clinal Variation in Populations of the Common Blue Butterfly Polyommatus icarus Rott. (Lepidoptera, Lycaenidae). Russ. J. Genet. 41, 859–870,
Brakefield, P. M., & Shreeve, T. (1992). Diversity within Populations. In The Ecology of Butterflies in Britain, edited by R H L Dennis, pp. 178-196. Oxford University Press.
Burghardt, F., Knüttel, H., Becker, M., & Fiedler, K. (2000). Flavonoid wing pigments increase attractiveness of female common blue (Polyommatus icarus) butterflies to mate-searching males. Naturwissenschaften, 87(7), 304-307.
Burghardt, F., Proksch, P., & Fiedler, K. (2001). Flavonoid sequestration by the common blue butterfly Polyommatus icarus: quantitative intraspecific variation in relation to larval hostplant, sex and body size. Biochemical Systematics and Ecology, 29(9), 875-889.
Kertész, K., Piszter, G., Horváth, Z. E., Bálint, Z., & Biró, L. P. (2017). Changes in structural and pigmentary colours in response to cold stress in Polyommatus icarus butterflies. Scientific Reports, 7(1), 1118.
Kertész, K., Piszter, G., Bálint, Z., & Biró, L. P. (2019). Biogeographical patterns in the structural blue of male Polyommatus icarus butterflies. Scientific reports, 9(1), 2338.
De Keyser, R., Breuker, C. J., Hails, R. S., Dennis, R. L., & Shreeve, T. G. (2015). Why small is beautiful: Wing colour is free from thermoregulatory constraint in the small lycaenid butterfly, Polyommatus icarus. PloS one, 10(4), e0122623.
Piszter, G., Bálint, Z., Kertész, K., Szatmári, L., Sramkó, G., & Biró, L. P. (2023). Breeding Polyommatus icarus Serves as a Large-Scale and Environmentally Friendly Source of Precisely Tuned Photonic Nanoarchitectures. Insects, 14(8), 716.
Sison-Mangus, M. P., Briscoe, A. D., Zaccardi, G., Knüttel, H., & Kelber, A. (2008). The lycaenid butterfly Polyommatus icarus uses a duplicated blue opsin to see green. Journal of Experimental Biology, 211(3), 361-369.

Magnificent shots! 🙂 And all we need to know in between. Thank you for sharing! Cheerz, UT
You’re welcome! Thank you 😊.
Such beautiful little jewels of butterflies and so much more complex than I could ever have imagined. Thank you for increasing my awareness.
You are welcome. Thank you for visiting!☺
The post was a highly informative.Nice work done.
Thank you!😊