
The upper wing surfaces of butterflies are often brightly coloured and visually highly apparent (above), whilst the undersides are usually fairly dull and inconspicuous (below). Although this characterisation is a gross generalisation – there are plenty of exceptions – these differences between the two wing surfaces, do reflect their contrasting functions: upper surfaces are used to signal messages – warnings aimed at predators, or sexual signals between males and females – whereas under surfaces, are generally a form of camouflage.

Let’s look at another example: the owl butterfly, or yellow-edged giant owl (Caligo atreus) to give it its full name (below). The underside, with its complex patterning is fairly cryptic, no doubt helping to camouflage this large butterfly, but look at the massive eyespot! Something else is going on: it is trying to look like an owl, or just very startling, surely aimed at frightening or freaking out anything thinking about trying to eat it! See it viewed upside down, here.

The upperside of the owl butterfly is a complete contrast: with extravagantly shiny blue (structural) and orange (pigmentary) colours (below). It only has to open its wings to transform itself from one state to the other. What a statement! These gorgeous upper surface colours have been called “seductive messages” by researcher Anupama Prakash, because they are aimed at trying to seduce the opposite sex, and we now know that this contrast occurs because certain genes are expressed on one side of the wing and not the other.


One rather obvious feature about butterfly wings, is that they are bilaterally symmetrical. Nothing too remarkable about that, so are we! The general idea, is that we can think of butterfly wings – at least for nymphalids – as having a sort of ancestral template, or prototype, called the nymphalid ground plan, or NGP for short (below).
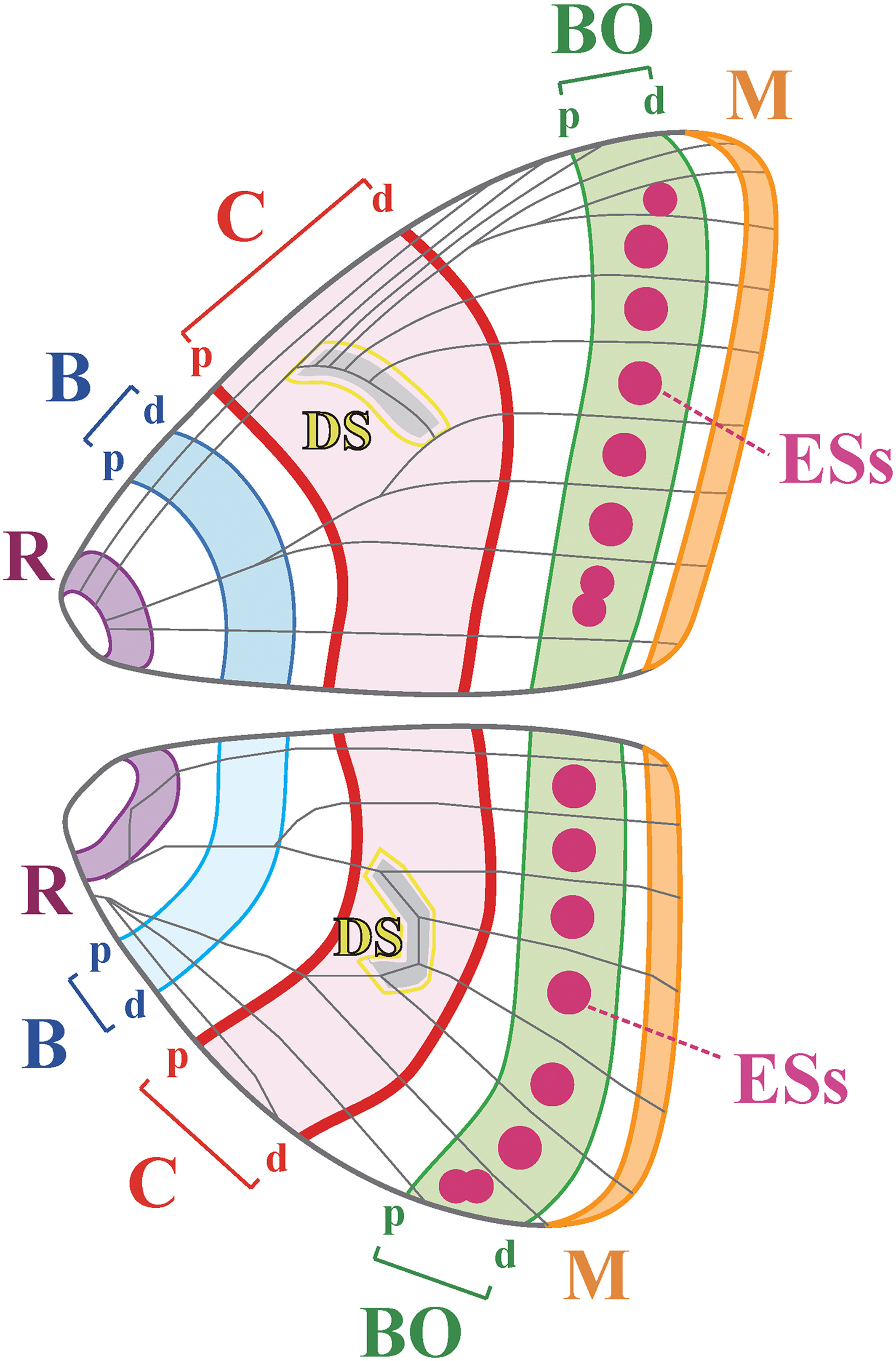
The NGP provides a conceptual framework to understand pattern variation in butterflies (Mazo-Vargas et al., 2017). There are in effect, autonomous colour patterns on the conceptual butterfly wing which are known as “symmetry systems” and scientists have been able to trace these compartments from one species to the next. In other words, there are homologous areas which are derived from a prototypical wing pattern. The diversity we see today is thought to have occurred as a result of evolution modifying the elements within these symmetry systems. The detail is more complicated than this, but the different colours and patterns have been assembled, over geological time, from these basic modules. In particular, from three general areas: the border, central, and basal symmetry systems (BO, C and B, above). There are also other components, like discal spots, eyespots and marginal bands, which can be tweaked into different sizes and shapes, by certain genes, over the course of evolution

Eyespots first appeared in nymphalid butterflies about 90 million years ago. At first, they were only present on the ventral surfaces of hindwings – where they have remained in many species, like the owl butterflies and morphos (below, last image), but over time, evolution has managed to co-opt them into appearing on the forewings as well; and more radically, they are also expressed on dorsal wing surfaces, where they mostly have a different function, to do with mate selection. In moving, they have changed their function, from helping the butterfly to survive, on the underside (under natural selection), to helping it find a mate, on the upperside (under sexual selection). But if that were not complicated enough, eyespots can have different functions on the same side. Look at this peacock pansy (Junonia almana) female (below). I would wager that the roundel like eyespots on the upper forewings – the ones that look like RAF aeroplane markings! – are involved in mate selection, whereas the ones on the hindwings (which can be flashed, like big eyes, by moving the forewings) are probably there to startle predators.

Different eyespot patterns are produced by altering the expression of regulatory and pigmentation genes. The formation of eyespot patterns is controlled by what is called a ‘developmental organizer’ (the focus), which induces the surrounding cells to synthesize specific pigments. The size of the eyespot is probably determined by the rate at which the chemical signal – a morphogen – causing the changes, diffuses outwards in a circle. In another butterfly, Bicyclus anynana, it has been determined that three genes are expressed in the centre of eyespots, in the black rings and in the surrounding gold rings. In other words, they control the formation of the rings of colour which go to make the eyespot.

Scientists seem to have had a lot of fun coming up with strange names for the genes, like spalt and hedgehog. Spalt genes determine the position of the wing veins in fruit flies, and are expressed in the centres of eyespots in butterflies. Hedgehog genes control segmentation in the embryo. The gene, Distal-less, seems to control the production of black (melanin) pigments within the eyespot, and also regulates their size. Variation in the expression of these genes, in the tiny (imaginal) wing discs growing inside the body of the caterpillar, affect the final size of the eyespots.

The so-called, apterous A gene, is involved in creating the different dorsal and ventral sides to a butterfly wing. If it is deleted, the butterfly will develop ventral (lower) surfaces patterns on both sides of its wings. I don’t suppose they were happy about that!
Certain proteins, called Wnt ligands, provide positional information as the tissues are developing and determine the position of symmetrical patterns and colours on the wing. optix and WntA genes have been called “paintbrush genes,” because they produce colourful patterns. Deleting the optix gene – which scientists can now do using ‘molecular scissors‘ – produces colourless butterflies; colourful species like the Painted lady, turn grey! In fact, the optix gene plays a fundamental role in the development of butterfly wings, generating colours and determining where they appear. In other cases however, deleting this gene generated new colours, that were not even present before!
Here is what the Common Jestor (Symbrenthia hippoclus) – see above – looks like on the upper or dorsal side.

During the course of evolution, butterflies have molded the basic elements of the nymphalid ground plan (see above) into a myriad different shapes, modifying the different components into various shapes and sizes, altering the colours, and swapping things about from underside to upperside, and from hindwings to forewings. Natural selection has modified a fairly limited number of genetic elements to generate incredibly diverse patterns; so that butterflies blend in with leaves, rock surfaces and tree bark, for example; or stand out so brightly, like blue Morpho wings (below), that they can be seen from passing aeroplanes!

The diversity of mimicry patterns found in Heliconius butterflies is also due to variation in just a handful of genes. For example, it is the optix gene, again, which controls the expression of red wing patterns in butterflies like the Doris longwing (Heliconius doris) (below).

Some of these cryptic or masquerade patterns – such as leaf-like patterns on the ventral sides of the wings of the Indian leaf butterfly, Kallima inachus (below), probably evolved gradually, passing through some early stages which were less than perfect, in terms of looking like a leaf, compared to the beautiful masquerade that was eventually arrived at and which we see today. Perhaps it can be further improved on by evolution?

The lovely calico patterns seen in butterflies in the genus Hamadryas (below), called ‘crackers’, appear to be an example of lichen crypsis. These butterflies – spend much of the day perched with their wings spread and pressed against the bark of lichen‐covered tree trunks and branches. Males use these positions as courtship territories and they produce a loud clicking noise, or crack, during flight.

Common graylings (below) have rock-like patterns – composed of minute ripples – on their under wing surfaces, which render this butterfly so well camouflaged when resting on trees or rocks. Black, brown and white colours intermingle in this complex pattern, but it is just possible to imagine how it might be produced by uneven gradients of colour-producing signals radiating out across a concentration gradient in the embryonic wing pad.

Finally, I will finish with some wonderful eyespots (below) on the undersides of a Blue morpho (Morpho peleides). I have tweaked the image a little to bring out the colours. We can think of the genes switching on and off during the development of the butterfly – whilst it was still a caterpillar – to produce these fantastic patterns!

References
Brunetti, C. R., Selegue, J. E., Monteiro, A., French, V., Brakefield, P. M., & Carroll, S. B. (2001). The generation and diversification of butterfly eyespot color patterns. Current Biology, 11(20), 1578-1585.
Martin, A., & Reed, R. D. (2014). Wnt signaling underlies evolution and development of the butterfly wing pattern symmetry systems. Developmental biology, 395(2), 367-378.
Mazo-Vargas, A., Concha, C., Livraghi, L., Massardo, D., Wallbank, R. W., Zhang, L., … & Breuker, C. J. (2017). Macroevolutionary shifts of WntA function potentiate butterfly wing-pattern diversity. Proceedings of the National Academy of Sciences, 114(40), 10701-10706.
Nijhout, H.F., 2017. The common developmental origin of eyespots and parafocal elements and a new model mechanism for color pattern formation. In Diversity and Evolution of Butterfly Wing Patterns (pp. 3-19). Springer, Singapore.
Monteiro, A. (2015). Origin, development, and evolution of butterfly eyespots. Annual review of entomology, 60, 253-271.
Prakash, A., & Monteiro, A. (2018). apterous A specifies dorsal wing patterns and sexual traits in butterflies. Proceedings of the Royal Society B: Biological Sciences, 285(1873), 20172685.
Suzuki, T. K., Tomita, S., & Sezutsu, H. (2014). Gradual and contingent evolutionary emergence of leaf mimicry in butterfly wing patterns. BMC evolutionary biology, 14(1), 229.
Suzuki, T. K., Tomita, S., & Sezutsu, H. (2019). Multicomponent structures in camouflage and mimicry in butterfly wing patterns. Journal of morphology, 280(1), 149-166.
Zhang, L., Mazo-Vargas, A. and Reed, R.D., 2017. Single master regulatory gene coordinates the evolution and development of butterfly color and iridescence. Proceedings of the National Academy of Sciences, 114(40), pp.10707-10712.

As usual fascinating reading Ray.
Thank you!